|
|
Artificial life |
|
|
Introduction to Genetic
Algorithms
|
|
Introduction to Genetic Algorithms. Physics, Biology, Economy or Sociology often have to deal with the classical problem of optimization. Economy particularly has become specialist of that field1. Generally speaking, a large part of mathematical development during the XVIIIth century dealt with that topic (remember those always repeated problems where you had to obtain the derivative of a function to find its extremes). Purely analytical methods widely proved their efficiency. They nevertheless suffer from a insurmountable weakness : Reality rarely obeys to those wonderful differentiable functions your professors used to show you2. Other methods, combining mathematical analysis and random search have appeared. Imagine you scatter small robots in a Mountainous landscape. Those robots can follow the steepest path they found. When a robot reaches a peak, it claims that it has found the optimum. This method is very efficient, but there's no proof that the optimum has been found, each robot can be blocked in a local optimum. This type of method only works with reduced search spaces. What could be the link between optimization methods and artificial life ? We are now 45 millions years ago examining a Basilosaurus :
Basilosaurus The Basilosaurus was quite a prototype of a whale. It was about 15 meters long for 5 tons. It still had a quasi-independent head and posterior paws. He moved using undulatory movements and hunted small preys3. Its anterior members were reduced to small flippers with an elbow articulation. Movements in such a viscous element (water) are very hard and require big efforts. People concerned must have enough energy to move and control its trajectory. The anterior members of basilosaurus were not really adapted to swimming4. To adapt them, a double phenomenon must occur : the shortening of the "arm" with the locking of the elbow articulation and the extension of the fingers which will constitute the base structure of the flipper.
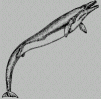
Tursiops flipper The image shows that two fingers of the common dolphin are hypertrophied to the detriment of the rest of the member. The basilosaurus was a hunter, he had to be fast and precise. Through time, subjects appeared with longer fingers and short arms. They could move faster and more precisely than before, and therefore, live longer and have many descendants. Meanwhile, other improvements occurred concerning the general aerodynamic like the integration of the head to the body, improvement of the profile, strengthening of the caudal fin ... finally producing a subject perfectly adapted to the constraints of an aqueous environment. This process of adaptation, this morphological optimization is so perfect that nowadays, the similarity between a shark, a dolphin or a submarine is striking. But the first is a cartilaginous fish (Chondrichtyen) originating in the Devonian (-400 million years), long before the apparition of the first mammal whose Cetacean descend from5. Darwinian mechanism hence generate an optimization process6, Hydrodynamic optimization for fishes and others marine animals, aerodynamic for pterodactyls, birds or bats. This observation is the basis of genetic algorithms. John Holland, from the University of Michigan began his work on genetic algorithms at the beginning of the 60s. A first achievement was the publication of Adaptation in Natural and Artificial System7 in 1975. Holland had a double aim : to improve the understanding of natural adaptation process, and to design artificial systems having properties similar to natural systems8. The basic idea is as follow : the genetic pool of a given population potentially contains the solution, or a better solution, to a given adaptive problem. This solution is not "active" because the genetic combination on which it relies is split between several subjects. Only the association of different genomes can lead to the solution. Simplistically speaking, we could by example consider that the shortening of the paw and the extension of the fingers of our basilosaurus are controlled by 2 "genes". No subject has such a genome, but during reproduction and crossover, new genetic combination occur and, finally, a subject can inherit a "good gene" from both parents : his paw is now a flipper. Holland method is especially effective because he not only considered the role of mutation (mutations improve very seldom the algorithms), but he also utilized genetic recombination, (crossover)9 : these recombination, the crossover of partial solutions greatly improve the capability of the algorithm to approach, and eventually find, the optimum. As an example, we're going to enter a world of simplified genetic. The "chromosomes" encode a group of linked features. "Genes" encode the activation or deactivation of a feature. Let us examine the global genetic pool of four basilosaurus belonging to this world. We will consider the "chromosomes" which encode the length of anterior members. The length of the "paw" and the length of the "fingers" are encoded by four genes : the first two encode the "paw" and the other two encode the fingers. In our representation of the genome, the circle on blue
background depict the activation of a feature, the cross on green
background depict its deactivation. The ideal genome (short paws
and long fingers) is : The genetic pool of our population is the following one :
We can notice that A and B are the closest to their ancestors ; they've got quite long paws and short fingers. On the contrary, D is close to the optimum, he just needs a small lengthening of his fingers. This is such a peculiar world that the ability to move is the main criteria of survival and reproduction. No female would easily accept to marry basilosaurus whose paws would look like A's. But they all dream to meet D one day. The fitness is easy to compute : we just have to give one point to each gene corresponding to the ideal. The perfect genome will then get four points. The probability of reproduction of a given subject will directly depend on this value. In our case, we'll get the following results :
We'll consider a cycle of reproduction with for descendants, i.e. four mating concerning height subjects. D will be selected four times and will then get four descendants. C will be selected twice and will get two descendants. Finally A and B will only be selected once. The reproduction pattern is the following :
During reproduction crossovers occur at a random place (center of the genome for A', B' and C', just after the first gene for D'). The link existing between the degree of adaptation and the probability of reproduction leads to a trend to the rise of the average fitness of the population. In our case, it jumps from 7 to 10. During the following cycle of reproduction, C' and D' will have a common descendant : D' : The new subject has inherited the intended genome : his paws have become flippers. We can then see that the principle of genetic algorithms is simple :
The functioning of a genetic algorithm can also be described in reference to genotype (GTYPE) and phenotype (PTYPE) notions10.
Crossover is the basis of genetic algorithms, there is nevertheless other operators like mutation. In fact, the desired solution may happen not to be present inside a given genetic pool, even a large one. Mutations allow the emergence of new genetic configurations which, by widening the pool improve the chances to find the optimal solution. Other operators like inversion are also possible, but we won't deal with them here. We saw before that in a genetic algorithm, the probability of reproduction directly depends on the fitness of each subject. We simulate that way the adaptive pressure of the environment. The use of this method nevertheless set two types of problems :
In order to palliate these problems, it's possible to transform the fitness values. Here are the four main methods : 1- Windowing : For each subject, reduce its fitness by the fitness of the worse subject. This permits to strengthen the strongest subject and to obtain a zero based distribution. 2- Exponential : This method, proposed by S.R. Ladd11, consists in taking the square roots of the fitness plus one. This permits to reduce the influence of the strongest subjects. 3- Linear Transformation : Apply a linear transformation to each fitness, i.e. f ' = a.f + b. The strongest subjects are once again reduced. 4- Linear normalization : Fitness are linearized. For example over a population of 10 subjects, the first will get 100, the second 90, 80 ... The last will get 10. You then avoid the constraint of direct reckoning. Even if the differences between the subjects are very strong, or weak, the difference between probabilities of reproduction only depends on the ranking of the subjects. To illustrate these methods, let's consider a population of four subjects to check the effect of scaling. For each subject, we give the fitness and the corresponding selection probability.
Windowing eliminates the weakest subject - the probability comes to zero - and stimulates the strongest ones (the best one jumps from 50 % to 67 %). Exponential flattens the distribution. It's very useful when a super-subject induces an excessively fast convergence. Linear transformation plays slightly the same role than exponential. At last, linear normalization is neutral towards the distribution of the fitness and only depends on the ranking. It avoids as well super-subjects as a too homogeneous distribution. Conclusion Genetic algorithms are original systems based on the supposed functioning of the Living12. The method is very different from classical optimization algorithms13.
It's important to understand that the functioning of such an algorithm does not guarantee success. We are in a stochastic system and a genetic pool may be too far from the solution, or for example, a too fast convergence may halt the process of evolution. These algorithms are nevertheless extremely efficient, and are used in fields as diverse as stock exchange, production scheduling or programming of assembly robots in the automotive industry. 1- The main paradigm of Economy (neo-classical) is largely just a wonderful ode to optimization mathematics. 2- That's what the recent "non linearity" revolution learned us. 3- S.J. Gould et al., Le livre de la vie, Seuil, Science ouverte, 1993, pp.186 ss. 4- It is said that basilosaurus reproduced on earth. In that case, posterior members were useful. Harrison R, Bryden M.M. dir., Baleines, dauphins et marsouins. Bordas, 1989. 5- This a very common phenomenon. In that case, we could also speak of the Ichthyosaure, marine reptile of the Mesozoic era whose morphology was closed to shark and dolphin. 6- It's important to understand that Darwinian process doesn't have to lead to optimum. It improves fitness but has nothing to do with optimum. 7- Holland J.H., Adaptation in natural and artificial system, Ann Arbor, The University of Michigan Press, 1975. 8- Goldberg D., Genetic Algorithms, Addison Wesley, 1988. 9- Emmeche C., Garden in the Machine. The Emerging Science of Artificial Life, Princeton University Press, 1994, pp. 114 ss. 10- Heudin J.C., La Vie Artificielle, Hermès, 1994, pp. 91 ss. 11- S.R. Ladd, Genetic Algorithm in C++, 1999-2000. Downloadable book. http://www.coyotegulch.com. 12- "Biological programming" is not limited to AG, another well-known case is neural networks. 13- Goldberg D, idem, pp. 8 ss. Jean-Philippe Rennard. Ph.D. May 2000 Copyright : This text has been written for an educational purpose. It is free for any private use. If you want to use it for a non-commercial public purpose, please quote author and source. Commercial use is strictly forbidden unless written agreement.
|
![]()

|
Copyright ©rennard.org. 2000, 2001, 2002.
|